
Функциональная единица мышечной ткани. Скелетные мышцы. Гладкая мышечная ткань нейрального происхождения
1. Виды мышечной ткани
2. Поперечно-полосатая скелетная ткань
6. Гладкая мышечная ткань
1. Свойством сократимости обладают практически все виды клеток, благодаря наличию в их цитоплазме сократительного аппарата, представленного сетью тонких микрофиламентов (5-7 нм), состоящих из сократительных белков - актина, миозина, тропомиозина и других. За счет взаимодействия названных белков микрофиламентов осуществляются сократительные процессы и обеспечивается движение в цитоплазме гиалоплазмы, органелл, вакуолей, образование псевдоподий и инвагинаций плазмолеммы, а также процессы фаго- и пиноцитоза, экзоцитоза, деления и перемещения клеток. Содержание сократительных элементов, а, следовательно, и сократительные процессы неодинаково выражены в разных типах клеток. Наиболее выражены сократительные структуры в клетках, основной функцией которых является сокращение. Такие клетки или их производные образуют мышечные ткани, которые обеспечивают сократительные процессы в полых внутренних органах и сосудах, перемещение частей тела относительно друг друга, поддержание позы и перемещение организма в пространстве. Помимо движения при сокращении выделяется большое количество тепла, а, следовательно, мышечные ткани участвуют в терморегуляции организма. Мышечные ткани неодинаковы по строению, источникам происхождения и иннервации, по функциональным особенностям. Наконец, следует отметить, что любая разновидность мышечной ткани, помимо сократительных элементов (мышечных клеток и мышечных волокон) включает в себя клеточные элементы и волокна рыхлой волокнистой соединительной ткани и сосуды, которые обеспечивают трофику мышечных элементов, осуществляют передачу усилий сокращения мышечных элементов на скелет. Однако, функционально ведущими элементами мышечных тканей являются мышечные клетки или мышечные волокна.
Классификация мышечных тканей
· Гладкая (неисчерченная)- мезенхимная;
· специальная - нейрального происхождения и эпидермального происхождения;
· Поперечно-полосатая (исчерченная)- скелетная;
· сердечная.
Как видно из представленной классификации мышечная ткань подразделяется по строению на две основные группы - гладкую и поперечно-полосатую. Каждая из двух групп в свою очередь подразделяется на разновидности, как по источникам происхождения, так и по строению и функциональным особенностям. Гладкая мышечная ткань , входящая в состав внутренних органов и сосудов, развивается из мезенхимы. К специальным мышечным тканям нейрального происхождения относятся гладкомышечные клетки радужной оболочки, эпидермального происхождения - миоэпителиальные клетки слюнных, слезных, потовых и молочных желез.
Поперечно-полосатая мышечная ткань подразделяется на скелетную и сердечную. Обе эти разновидности развиваются из мезодермы, но из разных ее частей: скелетная - из миотомов сомитов, сердечная - из висцерального листка спланхнотома.
Каждая разновидность мышечной ткани имеет свою структурно-функциональную единицу. Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и радужной оболочки является гладкомышечная клетка - миоцит; специальной мышечной ткани эпидермального происхождения - корзинчатый миоэпителиоцит ; сердечной мышечной ткани - кардиомиоцит ; скелетной мышечной ткани - мышечное волокно.
2. Поперечно-полосатая скелетная мышечная ткань
Структурно-функциональной единицей поперечно полосатой мышечной ткани является мышечное волокно . Оно представляет собой вытянутое цилиндрическое образование с заостренными концами длиной от 1 мм до 40 мм (а по некоторым данным до 120 мм), диаметром 0,1 мм. Мышечное волокно окружено оболочкой - сарколеммой, в которой под электронным микроскопом отчетливо выделяются два листка: внутренний - является типичной плазмолеммой, а наружный представляет собой тонкую соединительнотканную пластинку - базальную пластинку . В узкой щели между плазмолеммой и базальной пластинкой располагаются мелкие клетки - миосателлиты . Таким образом, мышечное волокно является комплексным образованием и состоит из следующих основных структурных компонентов :
· миосимпласта;
· клеток миосателиттов;
· базальной пластинки.
Базальная пластинка образована тонкими коллагеновыми и ретикулярными волокнами, относится к опорному аппарату и выполняет вспомогательную функцию передачи сил сокращения на соединительнотканные элементы мышцы.
Клетки миосателлиты являются камбиальными (ростковыми) элементами мышечных волокон и играют роль в процессах их физиологической и репаративной регенерации.
Миосимпласт является основным структурным компонентом мышечного волокна как по объему, так и по выполняемым функциям. Он образуется посредством слияния самостоятельных недифференцированных мышечных клеток - миобластов . Миосимпласт можно рассматривать как вытянутую гигантскую многоядерную клетку, состоящую из большого числа ядер, цитоплазмы (саркоплазмы), плазмолеммы, включений, общих и специальных органелл. В миосимпласте содержится несколько тысяч (до 10 000) продольно вытянутых светлых ядер, располагающихся на периферии под плазмолеммой. Вблизи ядер локализуются фрагменты слабовыраженной зернистой эндоплазматической сети, пластинчатого комплекса и небольшое число митохондрий. Центриоли в симпласте отсутствуют. В саркоплазме содержатся включения гликогена и миоглобина, аналога гемоглобина эритроцитов. Отличительной особенностью миосимпласта является также наличие в нем специализированных органелл , к которым относятся:
· миофибриллы;
· саркоплазматическая сеть;
· канальцы Т-системы.
Миофибриллы - сократительные элементы миосимпласта - в большом количестве (до 1000-2000) локализуются в центральной части саркоплазмы миосимпласта. Они объединяются в пучки, между которыми содержатся прослойки саркоплазмы. Между миофибриллами локализуется большое число митохондрий (саркосом). Каждая миофибрилла простирается продольно на протяжении всего миосимпласта и своими свободными концами прикрепляется к его плазмолемме у конических концов. Диаметр миофибриллы составляет 0,2-0,5 мкм. По своему строению миофибриллы неоднородны по протяжению и подразделяются на темные (анизотропные) или А-диски, и светлые (изотропные) или I-диски. Темные и светлые диски всех миофибрилл располагаются на одном уровне и обуславливают поперечную исчерченность всего мышечного волокна. Темные и светлые диски в свою очередь состоят из еще более тонких волоконец - протофибрилл или миофиламентов . Темные диски образованы более толстыми миофиламентами (10-12 нм), состоящими из белка миозина. Светлые диски образованы тонкими миофиламентами (5-7 нм), состоящими из белка актина. Посредине I-диска поперечно актиновым миофиламентам проходит темная полоска - телофрагма или Z-линия, посредине А-диска проходит менее выраженная М-линия или мезофрагма. Актиновые миофиламенты по средине I-диска скрепляются белками, составляющими Z-линию, свободными концами частично входит в А-диск между толстыми миофиламентами. При этом, вокруг одного миозинового филамента располагаются 6 актиновых. При частичном сокращении миофибриллы актиновые миофиламенты как бы втягиваются в А-диск и в нем образуется светлая зона или Н-полоска, ограниченная свободными концами актиновых миофиламентов. Ширина Н-полоски зависит от степени сокращения миофибриллы.
Участок миофибриллы, расположенный между двумя Z-линиями носит название саркомера и является структурно-функциональной единицей миофибриллы. Саркомер включает в себя А-диск и расположенные по сторонам от него две половины I-диска. Следовательно, каждая миофибрилла представляет собой совокупность саркомеров. Именно в саркомере осуществляется процесс сокращения. Следует отметить, что конечные саркомеры каждой миофибриллы прикрепляются к плазмолемме миосимпласта актиновыми миофиламентами. Структурные элементы саркомера в расслабленном состоянии можно выразить формулой :
Z+1/2I+1/2A+M+1/2A+1/2I+Z
Процесс сокращения осуществляется посредством взаимодействия актиновых и миозиновых филаментов и образования между ними актин-миозиновых мостиков , посредством которых происходит втягивание актиновых миофиламентов в А-диски укорочение саркомера. Для развития этого процесса необходимы три условия :
· наличие энергии в виде АТФ;
· наличие ионов кальция;
· наличие биопотенциала.
АТФ образуется в саркосомах (митохондриях) в большом числе локализованных между миофибриллами. Выполнение двух последних условий осуществляется с помощью еще двух специализированных органелл -саркоплазматической сети и Т-канальцев.
Саркоплазматическая сеть представляет собой видоизмененную гладкую эндоплазматическую сеть и состоит из расширенных полостей и анастомозирующих канальцев, окружающих миофибриллы. При этом саркоплазматическая сеть подразделяется на фрагменты, окружающие отдельные саркомеры. Каждый фрагмент состоит из двух терминальных цистерн, соединенных полыми анастомозирующими канальцами - L-канальцами. При этом терминальные цистерны охватывают саркомер в области I-дисков, а канальцы - в области А-диска. В терминальных цистернах и канальцах содержатся ионы кальция, которые при поступлении нервного импульса и достижении волны деполяризации мембран саркоплазматической сети, выходят из цистерн и канальцев и распределяются между актиновыми и миозиновыми миофиламентами, инициируя их взаимодействие. После прекращения волны деполяризации ионы кальция устремляются обратно в терминальные цистерны и канальцы. Таким образом, саркоплазматическая сеть является не только резервуаром для ионов кальция, но и играет роль кальциевого насоса.
Волна деполяризации передается на саркоплазматическую сеть от нервного окончания вначале по плазмолемме, а затем по Т-канальцам, которые не являются самостоятельными структурными элементами. Они представляют собой трубчатые выпячивания плазмолеммы в саркоплазму. Проникая вглубь, Т-канальцы разветвляются и охватывают каждую миофибриллу в пределах одного пучка строго на одном уровне, обычно на уровне Z-полоски или несколько медиальнее - в области соединения актиновых и миозиновых миофиламентов . Следовательно, к каждому саркомеру подходят и окружают его два Т-канальца. По сторонам от каждого Т-канальца располагаются две терминальные цистерны саркоплазматической сети соседних саркомеров, которые вместе с Т-канальцами составляют триаду. Между стенкой Т-канальца и стенками терминальных цистерн имеются контакты, через которые волна деполяризации передается на мембраны цистерн и обуславливает выход из них ионов кальция и начало сокращения. Таким образом, функциональная роль Т-канальцев заключается в передаче биопотенциала с плазмолеммы на саркоплазматическую сеть.
Для взаимодействия актиновых и миозиновых миофиламентов и последующего сокращения кроме ионов кальция необходима также энергия в виде АТФ, которая вырабатывается в саркосомах, в большом количестве располагающихся между миофибриллами.
Процесс взаимодействия актиновых и миозиновых филаментов упрощенно можно представить в следующем виде. Под влиянием ионов кальция стимулируется АТФ-азная активность миозина, что приводит к расщеплению АТФ, с образованием АДФ и энергии. Благодаря выделившейся энергии устанавливаются мостики между актином и миозином (а конкретнее, образуются мостики между головками белка миозина и определенными точками на актиновом филаменте)и за счет укорочения этих мостиков происходит подтягивание актиновых филаментов между миозиновыми. Затем эти связи распадаются (опять же с использованием энергии) и головки миозина образуют новые контакты с другими точками на актиновом филаменте, но расположенными дистальнее предыдущих. Так происходит постепенное втягивание актиновых филаментов между миозиновыми и укорочение саркомера . Степень этого сокращения зависит от концентрации ионов кальция вблизи миофиламентов и от содержания АТФ. После смерти организма АТФ в саркосомах не образуется, ее остатки расходуются на образование актин-миозиновых мостиков, а на распад уже не хватает, следствием чего наступает посмертное окоченение мышц, которое прекращается после аутолиза (распада) тканевых элементов.
При полном сокращении саркомера актиновые филаменты достигают М-полоски саркомера. При этом исчезают Н-полоски и I-диски, а формула саркомера может быть выражена в следующем виде:
Z+1/2IA+M+1/2AI+Z
При частичном сокращении формулу саркомера можно представить в следующем виде:
Z+1/nI+1/nIA+1/2H+M+1/2H+1/nAJ+1/nI+Z
Одновременное содружественное сокращение всех саркомеров каждой миофибриллы приводит к сокращению всего мышечного волокна. Крайние саркомеры каждой миофибриллы прикрепляются актиновыми миофиламентами к плазмолемме миосимпласта, которая на концах мышечного волокна имеет складчатый характер. При этом, на концах мышечного волокна базальная пластинка не заходит в складки плазмолеммы. Ее прободают тонкие коллагеновые и ретикулярные волокна, проникают в углубления складок плазмолеммы и прикрепляются в тех ее местах, к которым с внутренней стороны прикрепляются актиновые филаменты дистальных саркомеров. Благодаря этому создается прочная связь миосимпласта с волокнистыми структурами эндомизия. Коллагеновые и ретикулярные волокна концевых мышечных волокон, вместе с волокнистыми структурами эндомизия и перимизия в совокупности образуют сухожилия мышц, которые прикрепляются к определенным точкам скелета или вплетаются в сетчатый слой дермы в области лица. Благодаря сокращению мышц происходит перемещение частей или всего организма, а также изменение рельефа лица.
В мышечной ткани различают два основных типа мышечных волокон, между которыми имеются промежуточные, отличающиеся между собой прежде всего особенностями обменных процессов и функциональными свойствами и в меньшей степени - структурными особенностями.
Волокна I типа - красные мышечные волокна - характеризуются прежде всего высоким содержанием в саркоплазме миоглобина (что и придает им красный цвет), большим числом саркосом, высокой активностью в них сукцинатдегидрогеназы (СДГ), высокой активностью АТФ-азы медленного типа. Эти волокна обладают способностью медленного, но длительного тонического сокращения и малой утомляемостью.
Волокна II типа - белые мышечные волокна - характеризуются незначительным содержанием миоглобина, но высоким содержанием гликогена, высокой активностью фосфорилазы и АТФ-базы быстрого типа. Функционально характеризуются способностью быстрого, сильного, но непродолжительного сокращения. Между двумя крайними типами мышечных волокон находятся промежуточные, характеризующиеся различными сочетаниями названных включений и разной активностью перечисленных ферментов.
Мышца как орган состоит из мышечных волокон, волокнистой соединительной ткани, сосудов и нервов. Мышца - это анатомическое образование, основным и функционально ведущим структурным компонентом которого является мышечная ткань. Поэтому не следует рассматривать как синонимы понятия мышечная ткань и мышца.
Волокнистая соединительная ткань образует прослойки в мышце: эндомизий, перимизий и эпимизий, а также сухожилия. Эндомизий окружает каждое мышечное волокно, состоит из рыхлой волокнистой соединительной ткани и содержит кровеносные и лимфатические сосуды, в основном капилляры, посредством которых обеспечивается трофика волокна. Коллагеновые и ретикулярные волокна эндомизия проникают в базальную пластинку мышечного волокна, тесно с ним связаны и передают силы сокращения волокна на точки скелета. Перимизий окружает несколько мышечных волокон, собранных в пучки. В нем содержатся более крупные сосуды (артерии и вены, а также артериоло-венулярные анастомозы).
Эпимизий или фасция окружает всю мышцу, способствует функционированию мышцы, как органа. Любая мышца содержит все типы мышечных волокон в различном количественном соотношении. В мышцах, обеспечивающих поддержание позы, преобладают красные волокна, в мышцах, обеспечивающих движение пальцев и кистей, преобладают белые или переходные волокна. Характер мышечного волокна может меняться в зависимости от функциональной нагрузки и тренировки. Установлено, что биохимические, структурные и функциональные особенности мышечного волокна зависят от иннервации. Перекрестная пересадка эфферентных нервных волокон и их окончаний с красного волокна на белое и наоборот приводит к изменению обмена, а также структурных и функциональных особенностей в этих волокнах на противоположный тип.
3. Гистогенез и регенерация мышечной ткани
Из миотомов мезодермы в определенные участки мезенхимы выселяются малодифференцированные клетки - миобласты , часть из которых выстраивается в виде цепочки в стык друг к другу. В области контактов миобластов цитолеммы исчезает и образуется симпластическое образование - миотрубка, в которой ядра в виде цепочки располагаются в середине, а по периферии начинают дифференцироваться из миофиламентов миофибриллы. К миотрубке подрастают нервные волокна, образуя двигательные нервные окончания. Под влиянием эфферентной нервной импульсации начинается перестройка мышечной трубки в мышечное волокно: ядра перемещаются на периферию симпласта к плазмолемме, а миофибриллы занимают его центральную часть, из гладкой эндоплазматической сети развивается саркоплазматическая сеть, окружающая каждую миофибриллу на всем ее протяжении. Плазмолемма миосимпласта образует глубокие трубчатые впячивания - Т-канальцы. За счет деятельности зернистой эндоплазматической сети вначале миобластов, а затем и мышечных труб синтезируются и выделяются с помощью пластинчатого комплекса белки и полисахариды, из которых формируется базальная пластинка мышечного волокна.
Следует отметить, что при формировании миотрубки, а затем и дифференцировки мышечного волокна часть миобластов не входит в состав симпласта, а прилежит к нему, располагаясь под базальной пластинкой. Эти клетки носят название миосателлитов и играют важную роль в процессах физиологической и репаративной регенерации. Установлено, что закладка поперечно-полосатых скелетных мышечных волокон (миогенез) происходит только в эмбриональный период. В постнатальном периоде осуществляется их дальнейшая дифференцировка и гипертрофия, но количество мышечных волокон даже в условиях интенсивной тренировки не увеличивается.
Регенерация скелетной мышечной ткани
В мышечной, как в других тканях, различают два вида регенерации - физиологическую и репаративную. Физиологическая регенерация проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров. В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение. Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов. При определенных видах мышечной тренировки может формироваться преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров). Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1-2 года), что обусловлено прежде всего усилением нервной стимуляции. В старческом возрасте, а также в условиях малой мышечной нагрузки наступает атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку. Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуются мышечные почки , которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако, репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты). Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Советским ученым А. Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий :
· механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации;
· помещение измельченной ткани в фасциальное ложе;
· подшивание двигательного нервного волокна к измельченному трансплантату;
· наличие сократительных движений мышц-антагонистов и синергистов.
4. Иннервация и кровоснабжение скелетных мышц
Скелетные мышцы получают двигательную, чувствительную и трофическую (вегетативную) иннервацию. Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы - от двигательных нейронов определенных черепных нервов. При этом к каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется одним мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже сотни мышечных волокон получают двигательную иннервацию от одного мотонейрона, посредством разветвления его аксона.
Двигательное нервное волокно , подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обуславливая выход ионов кальция и начало процесса сокращения мышечного волокна.
Чувствительная (афферентная) иннервация скелетных мышц осуществляется псевдоуниполярными нейронами спинальных ганглиев, посредством разнообразных рецепторных окончаний дендритов этих клеток. Рецепторные окончания скелетных мышц можно разделить на две группы:
I. специфические рецепторные приборы, характерные только для скелетных мышц:
· мышечное веретено;
· сухожильный орган Гольджи;
II. неспецифические рецепторные окончания кустиковидной или древовидной формы, распределяющиеся в рыхлой соединительной ткани эндомизия, перимизия и эпимизия.
Мышечные веретена - довольно сложно устроенные инкапсулированные приборы. В каждой мышце содержится от нескольких единиц до нескольких десятков и даже сотен мышечных веретен. Каждое мышечное веретено содержит не только нервные элементы, но также 10-12 специфических мышечных волокон - интрафузальных , окруженных капсулой. Эти волокна располагаются параллельно сократительным мышечным волокнам (экстрафузальным) и получают не только чувствительную, но и специальную двигательную иннервацию. Мышечные веретена воспринимают раздражения как при растяжении данной мышцы, вызванном сокращением мышц-антагонистов, так и при ее сокращении.
Сухожильные органы представляют собой специализированные инкапсулированные рецепторы, включающие несколько сухожильных волокон, окруженных капсулой, среди которых распределяются терминальные ветвления дендрита псевдоуниполярного нейрона. При сокращении мышцы сухожильные волокна сближаются и сдавливают нервные окончания. Сухожильные органы воспринимают только степень сокращения данной мышцы. Посредством мышечных веретен и сухожильных органов при участии спинальных центров обеспечивается автоматизм движений (например, при ходьбе).
Трофическая иннервация обеспечивается вегетативной нервной системой (ее симпатической частью) и осуществляется в основном опосредованно, посредством иннервации сосудов.
Скелетные мышцы богато снабжаются кровью. В рыхлой соединительной ткани перимизия в большом количестве содержатся артерии и вены, артериолы, венулы и артериоло-венулярные анастомозы. В эндомизии располагаются только капилляры, преимущественно узкие (4,5-7 мкм), которые и обеспечивают трофику мышечного волокна. Мышечное волокно, вместе с окружающими его капиллярами и двигательным окончанием составляют мион. В мышцах содержится большое количество артериоло-венулярных анастомозов, обеспечивающих адекватное кровоснабжение при различной мышечной активности.
5. Сердечная поперечно-полосатая мышечная ткань
Структурно-функциональной единицей является клетка -кардиомиоцит . По строению и функциям кардиомиоциты подразделяются на две основные группы :
· типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
· атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50-120 мкм в длину, шириной 15-20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты , соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть - функциональный синтиций . Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (диском между кардиомиоцитами нет. Вставочные диски - это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы - аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов - атипичные кардиомиоциты образуют проводящую систему сердца , состоящую из:
· синусо-предсердный узел;
· предсердно-желудочковый узел;
· предсердно-желудочковый пучок (пучок Гиса)ствол, правую и левую ножки;
· концевые разветвления ножек - волокна Пункинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичным рядом особенностей :
· они крупнее (длина 100 мкм, толщина 50 мкм);
· в цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
· плазмолемма не образует Т-канальцев;
· во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности :
· Р-клетки (пейсмекеры) водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов - миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнееиз целомического эпителия этих участков.
Иннервация сердечной мышечной ткани
Биопотенциалы сократительные кардиомиоциты получают из двух источников:
· из проводящей системы сердца (прежде всего из синусо-предсердного узла);
· из вегетативной нервной системы (из ее симпатической и парасимпатической части).
Регенерация сердечной мышечной ткани
Кардиомиоциты регенерируют только по внутриклеточному типу. Пролиферации кардиомиоцитов не наблюдается. Камбиальные элементы в сердечной мышечной ткани отсутствуют. При поражении значительных участков миокарда (в частности, при инфаркте миокарда) восстановление дефекта происходит за счет разрастания соединительной ткани и образования рубцов (пластическая регенерация). Естественно, что сократительная функция в этих участках отсутствует. Поражение проводящей системы сопровождается нарушением ритма сердечных сокращений.
6. Гладкие мышечные ткани
Подавляющая часть гладкой мышечной ткани организма (внутренних органов и сосудов) имеет мезенхимальное происхождение.
Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и сосудов является миоцит . Представляет собой чаще всего веретенообразную клетку (длиной 20-500 мкм, диаметром 5-8 мкм), покрытую снаружи базальной пластинкой, но встречаются и отростчатые миоциты. В центре располагается вытянутое ядро, по полюсам которого локализуются общие органеллы: зернистая эндоплазматическая сеть, пластинчатый комплекс, митохондрии, цитоцентр. В цитоплазме содержатся толстые (17 нм) миозиновые и тонкие (7 нм) актиновые миофиламенты, которые располагаются в основном параллельно друг другу вдоль оси миоцита и не образуют А и I диски, чем и объясняется отсутствие поперечной исчерченности миоцитов. В цитоплазме миоцитов и на внутренней поверхности плазмолеммы встречаются многочисленные плотные тельца, к которым прикрепляются актиновые, миозиновые, а так же промежуточные филаменты. Плазмолемма образует небольшие углубления - кавеолы, которые рассматриваются как аналоги Т-канальцев. Под плазмолеммой локализуются многочисленные везикулы, которые вместе с тонкими канальцами цитоплазмы являются элементами саркоплазматической сети.
Механизм сокращения в миоцитах в принципе сходен с сокращением саркомеров в миофибриллах в скелетных мышечных волокнах. Он осуществляется за счет взаимодействия и скольжения актиновых миофиламентов вдоль миозиновых. Для такого взаимодействия также необходимы энергия в виде АТФ, ионы кальция и наличие биопотенциала. Биопотенциалы поступают от эфферентных окончаний вегетативных нервных волокон непосредственно на миоциты или опосредованно от соседних клеток через щелевидные контакты и передаются через кавеолы на элементы саркоплазматической сети, обуславливая выход из них ионов кальция в саркоплазму. Под влиянием ионов кальция развиваются механизмы взаимодействия между актиновыми и миозиновыми филаментами, аналогичные тем, которые происходят в саркомерах скелетных мышечных волокон, в результате чего происходит скольжение названных миофиламентов и перемещение плотных телец в цитоплазме. В миоцитах, кроме актиновых и миозиновых филаментов, имеются еще промежуточные, которые одним концом прикрепляются к цитоплазматическим плотным тельцам, а другим - прикрепительным тельцам на плазмолемме и таким образом передают усилия взаимодействия актиновых и миозиновых филаментов на сарколемму миоцита, чем и достигается его укорочение.
Миоциты окружены снаружи рыхлой волокнистой соединительной тканью - эндомизием и связаны друг с другом боковыми поверхностями. При этом, в области тесного контакта соседних миоцитов базальные пластинки прерываются. Миоциты соприкасаются непосредственно плазмолеммами и в этих местах имеются щелевидные контакты, через которые осуществляется ионная связь и передача биопотенциала с одного миоцита на другой, что приводит к одновременному и содружественному их сокращению. Цепь миоцитов, объединенных механической и метаболической связью, составляет функциональное мышечное волокно. В эндомизии проходят кровеносные капилляры, обеспечивающие трофику миоцитов, а в прослойках соединительной ткани между пучками и слоями миоцитов в перимизии проходят более крупные сосуды и нервы, а также сосудистые и нервные сплетения.
Эфферентная иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой. При этом, терминальные веточки аксонов эфферентных вегетативных нейронов, проходя по поверхности нескольких миоцитов, образуют на них небольшие варикозные утолщения, которые несколько прогибают плазмолемму и образуют мионевральные синапсы . При поступлении нервных импульсов в синаптическую щель выделяются медиаторы (ацетилхолин или норадреналин), и обуславливают деполяризацию мембран миоцитов и последующее их сокращение. Через щелевидные контакты биопотенциалы переходят из одного миоцита на другой, что сопровождается возбуждением и сокращением и тех гладкомышечных клеток, которые не содержат нервных окончаний. Возбуждение и сокращение миоцитов обычно продолжительны и обеспечивают тоническое сокращение гладкой мышечной ткани сосудов и полых внутренних органов, в том числе гладкомышечных сфинктеров. В этих органах содержатся и многочисленные рецепторные окончания в виде кустиков, деревцев или диффузных полей.
Регенерация гладкой мышечной ткани осуществляется несколькими способами:
· посредством внутриклеточной регенерациигипертрофии при усилении функциональной нагрузки;
· посредством митотического деления миоцитов при их повреждении (репаративная регенерация);
· посредством дифференцировки из камбиальных элементов - из адвентициальных клеток и миофибробластов.
7. Специальные гладкомышечные ткани
Нейрального происхождения развиваются из нейроэктодермы, из краев стенки глазного бокала, являющегося выпячиванием промежуточного мозга. Из этого источника развиваются миоциты, которые образуют две мышцы радужной оболочки глаза: мышцу суживающую зрачок и мышцу расширяющую зрачок. По своей морфологии миоциты радужной оболочки не отличаются от мезенхимных миоцитов, однако, отличаются по иннервации. Каждый миоцит получает вегетативную эфферентную иннервацию (мышца расширяющая зрачок - симпатическую, мышца суживающая зрачок -парасимпатическую). Благодаря этому, названные мышцы сокращаются быстро и координировано, в зависимости от мощности светового пучка.
Эпидермального происхождения развиваются из кожной эктодермы и представляют собой не типичные веретеновидные миоциты, а клетки звездчатой формы - миоэпителиальные клетки, располагающиеся в концевых отделах слюнных, молочных, слезных и потовых желез, снаружи от секреторных клеток. В своих отростках миоэпителиальные клетки содержат актиновые и миозиновые филаменты, благодаря взаимодействию которых отростки клеток сокращаются и способствуют выделению секрета из концевых отделов и мелких протоков названных желез в более крупные протоки. Эфферентную иннервацию получают также из вегетативного отдела нервной системы.
В статье мы рассмотрим виды мышечных тканей. Это очень важная тема в биологии, ведь каждый должен знать, как функционируют наши мышцы. Они представляют собой сложную систему, изучение которой, надеемся, вам будет интересно. А помогут лучше представить себе виды мышечной ткани картинки, которые вы найдете в этой статье. Прежде всего, дадим определение, которое необходимо при изучении данной темы.
Это особая группа и животных, основной функцией которой является ее сокращение, обусловливающее перемещение организма или составляющих его частей в пространстве. Данной функции соответствует строение основных элементов, из которых состоят различные виды мышечных тканей. Элементы эти имеют продольную и удлиненную ориентацию миофибрилл, включающих в свой состав - миозин и актин. Мышечная ткань, как и эпителиальная, это сборная тканевая группа, так как основные ее элементы развиваются из эмбриональных зачатков.
Сокращение мышечной ткани
Клетки ее, так же как и нервные, при воздействии электрических и химических импульсов могут возбуждаться. Способность их сокращаться (укорачиваться) в ответ на действие того или иного стимула связана с наличием миофибрилл, особых белковых структур, каждая из которых состоит из микрофиламентов, коротких белковых волокон. В свою очередь, они подразделяются на миозиновые (более толстые) и актиновые (тонкие) волокна. В ответ на нервное раздражение сокращаются различные виды мышечных тканей. Сокращение к мышце передается по нервному отростку через нейромедиатор, которым является ацетилхолин. Мышечные клетки в организме осуществляют энергосберегающие функции, так как расходуемая при сокращении различных мышц энергия выделяется затем в виде тепла. Именно поэтому, когда организм подвержен охлаждению, возникает дрожь. Это не что иное, как частые сокращения мышц.
Можно выделить следующие виды мышечных тканей, в зависимости от того, какое строение имеет сократительный аппарат: гладкую и поперечнополосатую. Они состоят из отличающихся по строению гистогенетических типов.
Мышечная ткань поперечнополосатая

Клетки миотомов, которые образуются из дорсальной мезодермы, являются источником ее развития. Эта ткань состоит из удлиненных имеющих вид цилиндров, концы которых заострены. 12 см в длину и 80 мкм в диаметре достигают эти образования. Симпласты (многоядерные образования) содержатся в центре мышечных волокон. Снаружи к ним прилегают клетки под названием "миосателлиты". Сарколеммой ограничены волокна. Она образуется плазмолеммой симпласт и базальной мембраной. Под базальной мембраной волокна располагаются миосателлиотоциты - так, что плазмолеммы симпласт касается их плазмолемма. Данные клетки являются камбиальным резервом мышечной скелетной ткани, и именно за счет него осуществляется регенерация волокон. Миосимпласты, кроме плазмолеммы, включают в себя также саркоплазму (цитоплазму) и расположенные по периферии многочисленные ядра.

Значение поперечнополосатой мышечной ткани
Описывая виды мышечной ткани, следует отметить, что поперечнополосатая является исполнительным аппаратом всей двигательной системы. Она формирует Кроме того, этот вид ткани входит в структуру внутренних органов, таких как глотка, язык, сердце, верхний отдел пищевода и др. Общая масса ее у взрослого человека составляет до 40% от массы тела, а у пожилых людей, а также новорожденных, ее доля - 20-30%.
Особенности поперечнополосатой мышечной ткани

Сокращение данного вида мышечной ткани, как правило, можно производить с участием сознания. Она обладает несколько большим быстродействием по сравнению с гладкой. Как вы видите, виды мышечной ткани отличаются (о гладкой мы поговорим совсем скоро и отметим некоторые другие различия между ними). В поперечнополосатых мышцах нервные окончания воспринимают информацию о текущем состоянии мышечной ткани, а затем передают ее по афферентным волокнам в нервные центры, ответственные за регуляцию двигательных систем. Управляющие сигналы поступают от регуляторов в виде нервных импульсов по двигательным или вегетативным эфферентным нервным волокнам.
Гладкая мышечная ткань

Продолжая описывать виды мышечных тканей человека, переходим к гладкой. Она формируется веретенообразными клетками, длина которых составляет от 15 до 500 мкм, а диаметр находится в промежутке от 2 до 10 мкм. В отличие от волокон мышцы поперечнополосатой, эти клетки имеют одно ядро. Кроме того, у них нет поперечной исчерченности.

Значение гладкой мышечной ткани
От сократительной функции этого вида мышечной ткани зависит функционирование всех систем организма, поскольку она входит в структуру каждой из них. Так, например, гладкая мышечная ткань участвует в управлении диаметром дыхательных путей, кровеносных сосудов, в сокращении матки, мочевого пузыря, в реализации двигательных функций нашего пищеварительного тракта. Она управляет диаметром зрачка глаз, а также участвует во множестве других функций различных систем организма.

Мышечные слои
Мышечные слои образует этот вид ткани в стенках лимфатических и кровеносных сосудов, а также всех полых органов. Обыкновенно это два или три слоя. Толстый циркулярный - наружный слой, средний присутствует не обязательно, тонкий продольный - внутренний. Питающие мышечную ткань кровеносные сосуды, а также нервы проходят параллельно оси мышечных клеток между их пучками. Гладкомышечные клетки можно разделить на 2 типа: унитарные (объединенные, сгруппированные) и автономные миоциты.
Автономные миоциты
Автономные функционируют довольно независимо друг от друга, так как нервным окончанием иннервируется каждая такая клетка. Они были обнаружены в мышечных слоях крупных кровеносных сосудов, а также в ресничной мышце глаза. Также к данному типу относятся клетки, из которых состоят мышцы, поднимающие волосы.
Унитарные миоциты
Унитарные мышечные клетки, напротив, тесно между собой переплетаются, так что мембраны их могут не просто примыкать плотно друг к другу, образуя десмосомы, но также и сливаться, формируя нексусы (щелевые контакты). Пучки образуются в результате данного объединения. Диаметр их составляет около 100 мкм, а длина достигает нескольких мм. Они формируют сеть, и в ее ячейки вплетаются Волокнами вегетативных нейронов иннервируются пучки, и они становятся функциональными единицами гладкой мышечной ткани. Деполяризация при возбуждении одной клетки пучка распространяется очень быстро на соседние, поскольку мало сопротивление щелевых контактов. Состоящие из унитарных клеток ткани есть в большинстве органов. К ним относятся мочеточники, матка, пищеварительный тракт.
Сокращение миоцитов
Сокращение миоцитов обусловлено в гладкой ткани, как и в поперечнополосатой, взаимодействием миозиновых и актиновых нитей. В этом схожи различные виды мышечной ткани у человека. Данные нити распределены внутри миоплазмы менее упорядоченно, чем в мышце поперечнополосатой. С этим связано отсутствие поперечной исчерченности в гладкой мышечной ткани. Внутриклеточный кальций является конечным исполнительным звеном, управляющим взаимодействием миозиновых и актиновых нитей (то есть сокращением миоцитов). Это же относится и к поперечнополосатой мышце. Однако детали механизма управления существенно отличаются от последней.
Проходящие в самой толще мышечной гладкой ткани вегетативные аксоны формируют не синапсы, что характерно для ткани поперечнополосатой, а многочисленные утолщения, имеющиеся по всей длине, которые и играют роль синапсов. Утолщения выделяют медиатор, который диффундирует к расположенным рядом миоцитам. Рецепторные молекулы находятся на поверхности этих миоцитов. С ними медиатор и взаимодействует. Он вызывает деполяризацию у миоцита внешней мембраны.
Особенности гладкой мышечной ткани
Нервная система, ее вегетативный отдел, управляется без участия сознания работой гладких мышц. Мышцы мочевого пузыря являются единственным исключением. Управляющие сигналы либо непосредственно реализуются, либо опосредованно - через гормональные (химические, гуморальные) воздействия.
Энергетические и механические свойства данного вида мышечной ткани обеспечивают поддержание тонуса (управляемого) стенок полых органов и сосудов. Связано это с тем, что гладкая ткань функционирует эффективно, не требуется больших затрат АТФ. У нее меньшее быстродействие, чем у мышечной ткани поперечнополосатой, однако она способна сокращаться более продолжительное время, кроме того, может развивать существенное напряжение и изменять в широких пределах свою длину.
Итак, мы рассмотрели виды мышечных тканей и особенности их структурной организации. Конечно, это лишь основная информация. Можно долго описывать виды мышечных тканей. Рисунки помогут вам наглядно их представить.
Структурно-функциональной единицей поперечнополосатой скелетной мышечной ткани является мышечное волокно. Волокно может достигать 12 см в длину, содержит большой объем саркоплазмы и сотни ядер. Каждое волокно покрыто сарколеммой, состоящей из двух слоев: внутреннего - плазмолеммы толщиной 8-10 нм и внешнего - базальной мембраны толщиной 30-40 нм. Между плазмолеммой и базальной мембраной имеется пространство шириной 15-25 нм. Кроме того, в базальную мембрану вплетаются ретикулярные волокна.
Значительный объем саркоплазмы занимают сократительные органеллы - миофибриллы. Каждая миофибрилла состоит из большого числа правильно чередующихся темных и светлых полос (дисков). В поляризованном свете темные диски обнаруживают двойное лучепреломление, поэтому называются анизотропными (А-дисками). Светлые диски таким свойством не обладают и называются изотропными (I-дисками). Каждая миофибрилла образована пучком параллельно идущих миофиламентов. А-диски состоят из толстых и тонких миофиламентов, а I-диски - только из тонких. Тонкие филаменты (5-8 нм) образованы белками актином, тропомиозином, тропонином, а толстые (10-12 нм) - миозином, белками М- и Н-полос и другими. Тонкие филаменты располагаются между толстыми, образуя гексагональное расположение.
Структурно-функциональной единицей миофибриллы является саркомер . Условная формула саркомера - 1/2 1-диска + А-диск + 1/2 I-диска. Линия сшивки соседних саркомеров соответствует Z-линии (телофрагме), которая состоит из белков альфа-актинина, десмина, вимен-тина. У позвоночных длина саркомера равна 2-3 мкм. Средняя часть миозинового диска, куда не доходят актиновые миофиламен-ты, более светлая и называется Н-полоской. Ее пересекает М-линия (мезофрагма), скрепляющая миозиновые нити посередине саркомера. В подмембранном слое сим-пласта обнаружены белки винкулин и спектрин, входящие в состав скелета симпласта.
Компоненты метаболической среды симпласта хорошо выражены. В гистогенезе с возрастанием степени зрелости симпластов наблюдается увеличение числа митохондрии, которые ориентируются по бокам Z-линии между миофибриллами и под сарколеммой. Гранулы гликогена, липидные капли формируют скопления между миофибриллами и под сарколеммой. Содержание миоглобина (связывающий кислород пигмент) варьирует в зависимости от образа жизни животного. Рибосомы представлены в виде полисом. Небольшое число лизосом принимают участие в процессах внутрисимпластической регенерации. Клеточный центр в симпласте отсутствует.
Саркоплазматическая сеть и Т-трубочки развиваются параллельно. Последние - это инвагинации плазмолеммы, которые опоясывают каждый саркомер. В продольном направлении вокруг каждой миофибриллы идут канальцы саркоплаз-матической сети. Так формируются продольная и поперечная системы, которые на срезах видны как триады. Триада - это комплекс, состоящий из поперечной трубочки и профилей двух цистерн саркоплазматической сети, расположенных симметрично по обе стороны от Т-трубочки. В цистернах саркоплазматической сети накапливаются ионы кальция, необходимые для сокращения миофибрилл.
В позднем онтогенезе происходит ряд ультраструктурных изменений в клетках и симпластах. Наиболее значимы - утолщение базальной мембраны, дезорганизация миофибрилл и Z-линии, возникновение скоплений митохондрий под сарколеммой, отделение миосателлитоцитов от симпласта и переход их в интерстициальное пространство. Иннервация мышечных волокон осуществляется двигательными нейронами передних рогов спинного мозга, которые формируют нервно-мышечные синапсы примерно в центральной части волокна.
Регенерация . Для успешной регенерации мышечной ткани необходимо сохранение напряжения мышцы, восстановление кровоснабжения и нервной связи. Основным источником регенерации являются миосателлитоциты. После активации последних происходит их митотическое деление, возникают миобласты, которые претерпевают дифференцировку, сливаются друг с другом и формируют симпласты. Развитие симпластов продолжается с участием размножающихся миосателлитоциов, часть которых сливается с растущими симпластами. Так формируются новые клеточно-симпластические системы - мышечные волокна.
Цели занятия. Научиться:
1.Определять на светооптическом уровне исчерченную, неисчер-ченную(гладкую),сердечную мышечные ткани.
2.Узнавать и анализировать отличия разных видов мышечной ткани.
3.Анализировать на электронно-оптическом уровне структуру мышечного волокна, гладкой мышечной клетки, кардиомиоцитов.
1.Что является источником формирования исчерченной мускулатуры органов опоры и движения?
2. Каково функциональное назначение скелетных мышц?
3. Принципы работы мышц.
4.Строение скелетной мышцы как органа.
5.Из каких мышц построены стенки полых внутренних органов?
Имея разное происхождение и строение, мышечные ткани объединяет способность к сокращению. Сократительный аппарат занимает значительную часть в цитоплазме, в его составе присутствуют актиновые и миозиновые филаменты, которые формируют органеллы специального значения–миофибриллы.
По морфофункциональному признаку различают:
· Скелетную или поперечно-полосатую или исчерченную мышечную ткань . Начало и прикрепление мышц находится на скелете. Мышцы являются произвольными, поскольку их сокращения подчиняются нашей воле. К этой группе мышц относят скелетные мышцы, мышцы языка, гортани и др.
· Сердечная мышечная ткань входит в состав мышечной стенки сердца. Иннервируется вегетативными нервами, является непроизвольной.
· Гладкая (неисчерченная) мышечная ткань характеризуется отсутствием исчерченности, а поскольку сокращения также не подчиняются нашей воле, то мышцы называют непроизвольными. Иннервация осуществляется вегетативной нервной системой. Из гладких мышц построены стенки внутренних органов, стенка сосудов.
В зависимости от источников развития выделяют пять типов мышечной ткани:
1. Мезенхимного происхождения (гладкая мышечная ткань).
2.Из кожной эктодермы и прехордальной пластинки – миоэпителиальные клетки (например, в потовых, слюнных железах).
3.Нейральное происхождение (из нервной трубки)–мышцы суживающие и расширяющие зрачок.
4.Целомическое происхождение (миоэпикардиальная пластинка) – сердечная мышечная ткань.
5. Из миотомов мезодермы– исчерченная мышечная ткань.
Исчерченная (поперечно-полосатая) мышечная ткань Источником развития являются клетки миотомов миобласты. Различают головные, шейные, грудные, поясничные, крестцовые миотомы. Они разрастаются в дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых нервов.
Часть миобластов дифференцируется на месте (образуют аутохтонную мускулатуру), а другие, с 3 недели внутриутробного развития мигрируют в мезенхиму и сливаясь, друг с другом образуют мышечные трубки (миотубы) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра смещаются к периферии. Клеточные центры и микротрубочки исчезают. Гранулярная ЭПС значительно редуцируется. Такая многоядерная структура называется симпласт, а для мышечной ткани - миосимпласт.
Часть миобластов дифференцируется в миосателлитоциты, которые располагаются на поверхности миосимпластов и принимают участие в регенерации мышечной ткани.
Структурной единицей мышечной ткани является мышечное волокно , состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной (рис.21). Длина мышечных волокон колеблется от 1 до 40 мм, а толщина 0,1 мм.
В мышечном волокне различают мембранный аппарат, фибриллярный (сократительный) аппарат, трофический аппарат (ядро, саркоплазма, цитоплазматические органеллы).
Мембранный аппарат . Каждое мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану и плазмолемму (под базальной мембраной), плазмолемма образует впячивания (Т- трубочки).
К плазмолемме снаружи прилежат миосателлитоциты. При повреждении базальной мембраны запускается митотический цикл миосателлитоцитов.
Фибриллярный аппарат .Исчерченные волокна можно разделить на составляющие их фибриллы (диаметром 1 мкм), названные миофибриллами. В мышечном волокне они ориентированы продольно.
При рассматривании мышечных волокон и миофибрилл под световым микроскопом, отмечается чередование в них темных и светлых участков – дисков. Темные диски отличаются двойным лучепреломлением и называются анизотропными дисками или А- дисками. Светлые диски не обладают двойным лучепреломлением и называются изотропными или I – дисками. В средней части диска А имеется более светлый участок Н-зона (участок содержащий только толстые нити белка миозина). В области Н-зоны выделяется более темная М-линия, состоящая из миомезина (необходим для сборки толстых нитей и их фиксации при сокращении). В середине диска I расположена плотная линия Z, которая построена из белковых фибриллярных молекул. В частности, большую роль играет альфа актинин. Z – линия соединена с соседними миофибриллами с помощью белка десмина и поэтому все названные линии и диски соседних миофибрилл совпадают и создается картина поперечно-полосатой исчерченности мышечного волокна.
Структурной единицей миофибриллы является саркомер (S) - это пучок миофиламентов заключенный между двумя Z линиями (рис.22). Принимая во внимание вышеуказанные обозначения можно структуру саркомера записать в виде формулы:
S= Z 1 + 1/2 I 1 + А + 1/2 I 2 + Z 2
Под электронным микроскопом миофибриллы представляют агрегаты из толстых (меозиновых) филаментов (диаметр 14 нм, длина 1500 нм, расстояние между ними 20-30 нм). Между толстыми филаментами располагаются тонкие филаменты (диаметр 7-8 нм).
Толстые филаменты (миозиновые нити) состоят из молекул белка миозина. Он является важнейшим сократительным белком мышцы. При непосредственном участии миозина химическая энергия трансформируется в механическую работу. Каждая миозиновая нить состоит из 300-400 молекул миозина. Молекула миозина – это гексамер, состоящий из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две спирально закрученные полипептидные нити. Они несут на своих концах глобулярные (шаровидные) головки. Между головкой и тяжелой цепью – шарнирный участок, с помощью которого головка может изменять свою конфигурацию. В области головок - легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким образом, что их головки обращены наружу, выступая над поверхностью толстой нити, а тяжелые цепи образуют стержень толстой нити.
Тяжелые и легкие цепи в молекуле миозина можно разделить обработкой мочевиной, гуанидинхлоридом и др. При мягкой обработке можно отделить только легкие цепи. Миозину свойственна АТФ-азная активность – высвобождающаяся энергия используется для мышечного сокращения.
Тонкие нити (актиновые нити). Образованы тремя белками: актином, тропонином и тропомиозином. Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы тропонина располагаются вдоль спирали.
Толстые нити занимают центральную часть саркомера–А-диск, тонкие занимают I диски и частично входят между толстыми миофиламентами. Только толстые нити содержит Н-зона.
При поступлении нервных импульсов по аксонам двигательных нейронов происходит сокращение мышечного волокна . Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка) и окружено сетью гемокапилляров, располагающихся в прилежащей рыхлой соединительной ткани. Этот комплекс называется мион. Группа мышечных волокон, которые иннервируются одним мотонейроном называется нервно-мышечной единицей. Мышечные волокна в этом случае могут располагаться не рядом (одно нервное окончание может контролировать от одного до десятков мышечных волокон).
В покое взаимодействие тонких и толстых нитей (миофиламентов) невозможно, т.к. миозин-связывающие участки актина заблокированы тропомиозином. При высокой концентрации ионов кальция конформационные (пространственные) изменения тропомиозина приводят к разблокированию миозин-связывающих участков молекул актина. Плазмолемма миосимпласта образует пальцевидные впячивания (инвагинации) ориентированные поперечно по отношению к миосимпласту называемые Т-трубочки. К каждой Т-трубочке примыкают по две цистерны саркоплазматического ретикулума (гладкая ЭПС), образуя триаду: две цистерны и одна Т-трубочка. Са 2+ концентрируется в цистернах (там его концентрация в 800 раз больше, чем в саркоплазме).
Механизм сокращения.При поступлении нервного импульса волна деполяризации доходит до цистерн саркоплазматического ретикулума, из них выделяются ионы кальция и концентрация кальция в саркоплазме резко возрастает. Са 2+ диффундирует к тонким нитям (филаментам) саркомера, где связывается с тропонином и миозиновыми головками. Это приводит:
1.К изменению конформации (пространственного расположения) тропомиозина, что, в свою очередь, приводит к освобождению участков актина, необходимых для взаимодействия с миозиновыми головками.
2.Появлению АТФ-азной активности миозина.
3.Взаимодействию миозиновых головок с актином (актино- миозиновые «мостики»).
Все это вместе взятое приводит к тому, что миозиновые головки «шагают» по актину, образуя в ходе перемещения новые связи актина и миозина, сближая две Z-линии. При сокращении уменьшаются только светлые диски.
Расслабление. Са 2+ -АТФ-аза саркоплазматического ретикулума закачивает Са 2+ из саркоплазмы в цистерны. В саркоплазме концентрация Са 2+ становится низкой. Са 2+ -тропомиозин закрывает миозин-связывающие участки тонких нитей и препятствует их взаимодействию с миозином.
Чувствительная иннервация (нервно-мышечные веретена). Интрафузальные мышечные волокна вместе с чувствительными нервными окончаниями формируют нервно-мышечные веретена, являющиеся рецепторами скелетной мышцы. Снаружи сформирована капсула веретена. При сокращении поперечно-полосатых (исчерченных) мышечных волокон изменяется натяжение соединительно-тканной капсулы веретена и соответственно изменяется тонус интрафузальных (расположенных под капсулой) мышечных волокон. Формируется нервный импульс.
Классификация и типы мышечных волокон. Скелетные мышцы, состоящие из мышечных волокон отличаются по многим параметрам: цвету, диаметру, утомляемости, скорости сокращения и т.д. В каждой мышце присутствуют разные типы мышечных волокон. В исчерченных мышцах различают два вида мышечных волокон: экстрафузальные, которые преобладают и обуславливают собственно сократительную функцию мышцы и интрафузальные, входящие в состав проприоцепторов–нервно-мышечных веретен.
По характеру сокращения мышечные волокона делят на фазные и тонические . Фазные способны осуществлять быстрые сокращения, но не могут длительно удерживать достигнутый уровень укорочения. Тонические –обеспечивают поддержание статического напряжения или тонуса.
По биохимическим особенностям и цвету выделяют красные и белые мышечные волокна . Цвет мышцы обусловлен степенью васкуляризации. Кроме того, существует прямая корреляция между содержанием миоглобина и цветом мышцы. Характерной особенностью красных мышечных волокон является наличие многочисленных митохондрий, цепи которых располагаются между миофибриллами. В белых мышечных волокнах митохондрий меньше и они располагаются равномерно в саркоплазме мышечного волокна.
Скорость сокращения определяется типом миозина . Высокую скорость сокращения обеспечивает быстрый миозин (для него характерна высокая активность АТФ-азы); меньшая скорость сокращения характерна для медленного миозина (характерна невысокая активность АТФ-азы). Следовательно, по активности АТФ-азы можно судить и о наборе миозинов.
Тип окислительного обмена . Мышечные волокна используют два пути образования АТФ:
* при анаэробном типе метаболизма из 1 молекулы глюкозы образуется 2 молекулы АТФ и молочная кислота.
* при аэробном окислении из 1 молекулы глюкозы образуется 38 молекул АТФ и конечные продукты метаболизма: СО 2 и Н 2 О. Идентификация мышечных волокон основана на выявлении активности фермента сукцинатдегидрогеназы (СДГ), которая является маркером для митохондрий и цикла Кребса. Активность этого фермента свидетельствует о напряженности энергетического метаболизма. Выделяют мышечные волокна А-типа (гликолитические) с низкой активностью СДГ, С-тип (оксидативные) с высокой активностью СДГ. Мышечные волокна В-типа занимают промежуточное положение. Переход мышечных волоко от А-типа в С-тип маркирует изменения от анаеробного гликолиза к метаболизму, зависящему от кислорода.
Существует много и других классификаций.
Факторами, определяющими структуру и функцию скелетной мышцы являются влияние нервной ткани, гормональное влияние, уровень васкуляризации, уровень двигательной активности и местоположение мышцы.
Сердечная мышечная ткань находится в мышечной оболочке сердца (миокард) и в устьях связанных с ним крупных сосудов. Имеет клеточный тип строения и основным функциональным свойством служит способность к спонтанным ритмическим сокращениям.
Развивается из миоэпикардиальной пластинки (висцеральный листок спланхнотома в шейном отделе), клетки которой размножаются митозом а потом дифференцируются. В клетках появляются миофиламенты, которые далее формируют миофибриллы.
Сердечная мышечная ткань образована клетками, которые называются кардиомиоциты. Между ними расположена рыхлая соединительная ткань и кровеносные сосуды с помощью вставочных дисков. Кардиомиоциты обьединяются в мышечные «волокна». Продольные и боковые связи кардиомиоцитов обеспечивают функциональное единство миокарда. Последние являются комплексом контактов. На поперечном срезе вставочных дисков выявляют десмосомы и щелевидные контакты (нексусы).
Выделяют рабочие (сократительные) кардиомиоциты , которые образуют цепочки клеток и обеспечивают силу сокращения сердечной мышцы. Клетки удлиненной формы с центрально расположенным ядром (рис.23). Вблизи ядра (или двух) комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Имеются Т-трубочки и L-трубочки. Десмосомы обеспечивают механическое сцепление, которое препятствует расхождению кардиомиоцитов. Щелевидные контакты способствует передаче сокращения от одного кардиомиоцита к другому.
Проводящие (атипичные) кардиомиоциты –среди них различают: 1.Водители ритма–это клетки небольших размеров, в саркоплазме мало гликогена, мало миофибрилл и они расположены по периферии. Клетки имеют хорошее кровоснабжение и иннервацию. Они воспринимают сигналы от нервных окончаний и способны автоматически генерировать сигналы обеспечивающие ритмические сокращения сердца.
2.Проводящие (переходные) кардиомиоциты проводят возбуждение от водителя ритма. Образуют длинные волокна. Миофибриллы в небольшом количестве, имеют спиральный ход, мелкие митохондрии, немного гликогена.
3.Волокна Пуркинье–являются самыми крупными клетками в мышечной ткани сердца с неупорядоченным расположением миофибрилл, множеством мелких митохондрий, много гликогена, нет Т-трубочек, клетки связаны между собой десмосомами и щелевидными контактами.
Секреторные кардиомиоциты – находятся в, основном, в предсердиях, преимущественно в правом. Характеризуются отростчатой формой и слабым развитием сократительного аппарата. В саркоплазме, вблизи полюсов ядра-секреторные гранулы, содержащие атриопептин (гормон, регулирующий артериальное давление). Гормон вызывает потерю натрия и воды с мочой, расширение сосудов, снижение давления, угнетение секреции альдостерона, кортизола, вазопрессина.
Сократительный аппарат рабочих кардиомиоцитов сходен со скелетными мышечными волокнами. Миофибриллы в кардиомиоците могут объединяться в комплексы, образуя единые сократительные структуры. В саркоплазме миофибриллы ориентированы продольно и располагаются преимущественно по периферии. Саркотубулярная система вцелом имеет сходство с исчерченными мышечными волокнами. Саркоплазматическая сеть развита слабее, не так активно накапливает Са 2. . При расслаблении, ионы кальция выделяются в саркоплазму с низкой скоростью, что обеспечивает автоматизм и частые сокращения кардиомиоцитов. Т-трубочки широкие и образуют диады (одна Т-трубочка и одна цистерна сети), которые сходятся в области Z-линии. Энергетический аппарат-это митохондрии и включения.
Неисчерченная (гладкая) мышечная ткань. Структурно-функциональной единицей данной ткани являются гладкие мышечные клетки (ГМК), которые способны к гипертрофии и регенерации. Они образуют стенки внутренних полых органов, сосудов. Более крупные по размерам ГМК характерны для стенок полых внутренних органов, а меньших размеров–для стенки сосудов. Клетки контролируют моторику, величину просвета. Имеют веретенообразную форму, в центре палочковидной формы ядро. В ГМК отсутствует поперечно-полосатая исчерченность. ГМК окружены сарколеммой, которая снаружи покрыта базальной мембраной. Длина от 20 мкм до 1 мм. В саркоплазме у полюсов находится комплекс Гольджи, много митохондрий, рибосом, развит саркоплазматический ретикулум. Миофиламенты расположены вдоль продольной оси. В ГМК актиновые и миозиновые филаменты не формируют миофибрилл. Актиновые нити (тонкие филаменты) ориентированы по продольной оси ГМК. По количеству их больше и они прикрепляются к плотным тельцам, которые являются специальными сшивающими белками. Рядом с актиновыми нитями располагаются мономеры миозина (микромиозин). Обладая разной длиной они, значительно короче тонких нитей.
Сокращение гладких мышечных клеток осуществляется при взаимодействии актиновых филаментов и миозина. Сигнал идущий по нервным волокнам обуславливает выделение медиатора, что изменяет состояние сарколеммы. Она образует колбовидные впячивания (кавеолы), где концентрируются ионы кальция. Сокращение ГМК индуцируется притоком ионов кальция в саркоплазму (кавеолы отшнуровываются и вместе с ионами кальция попадают в саркоплазму). Это приводит к полимеризации миозина и взаимодействию его с актином. Актиновые нити и плотные тельца сближаются, усилие передается на сарколемму и ГМК укорачивается. Миозин ГМК способен взаимодействовать с актином только после фосфорилирования его легких цепей особым ферментом–киназой легких цепей. После прекращения сигнала ионы кальция покидают кавеолы; миозин деполяризуется, теряет сродство к актину. В результате комплексы миофиламентов распадаются; сокращение прекращается.
Особые типы гладких мышечных клеток. Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности. Окружают секреторные отделы и выводные протоки желез (слюнных, молочных, слезных). С железистыми клетками они связаны десмосомами. Сокращаясь, способствуют выделению секрета. В концевых (секреторных) отделах форма клеток отросчатая, звездчатая. Ядро в центре, в цитоплазме, преимущественно в отростках локализованы миофиламенты, которые образуют сократительный аппарат. В этих клетках есть и цитокератиновые промежуточные филаменты, что подчеркивает их сходство с эпителиоцитами.
Мионейральные клетки – развиваются из клеток наружного слоя глазного бокала и образуют мышцу, суживающую зрачок и мышцу, расширяющую зрачок. По строению первая мышца сходна с ГМК мезенхимного происхождения. Мышца, расширяющая зрачок образована отростками клеток, располагающимися радиально, а ядросодержащая часть клетки находится между пигментным эпителием и стромой радужки.
Миофибробласты относятся к рыхлой соединительной ткани и представляют собой видоизмененные фибробласты. Они проявляют свойства как фибробластов, так и ГМК (обладают выраженными сократительными свойствами). Как вариант этих клеток можно рассматривать миоидные клетки в составе стенки извитого семенного канальца яичка и наружного слоя теки фолликула яичника. При заживлении раны часть фибробластов синтезирует гладкомышечные актины и миозины.
Эндокринные гладкие миоциты –это видоизмененные ГМК, представляющие основной компонент юкста-гломерулярного аппарата почек. Они находятся в стенке артериол почечного тельца, имеют хорошо развитый синтетический аппарат и редуцированный сократительный. Продуцируют фермент ренин, находящийся в гранулах и попадающий в кровь механизмом экзоцитоза.
Вопросы для самоконтроля :
1.Как классифицируются мышечные ткани по морфо-функциональному признаку? по происхождению?
2.Что является структурно-функциональной единицей мышечной ткани?
3.Строение фибриллярного аппарата мышечных волокон.
4.Напишите формулу саркомера.
5.Строение мышечных волокон под световым и электронным микроскопом.
6.Механизм сокращения и расслабления мышечного волокна.
7.Как классифицируются мышечные волокна? Типы мышечных волокон?
8.Виды кардиомиоцитов в сердечной мышце, особенности их строения.
9.Строение ГМК.
10.Перечислить особые типы гладких мышечных клеток.
Конец работы -
Эта тема принадлежит разделу:
Общая гистология
Кафедра гистологии эмбриологии и цитологии.. общая гистология ижевск..
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
Мышечные ткани представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака - выраженной сократительной способности, благодаря которой они могут выполнять свою основную функцию - перемещать тело или его части в пространстве.
Важнейшие свойства мышечных тканей. Структурные элементы мышечных тканей (клетки, волокна) обладают удлиненной формой и способны к сокращению благодаря мощному развитию сократительного аппарата. Для последнего характерно высокоупорядоченное расположение актиновых и миозиновых миофиламентов, создающее оптимальные условия для их взаимодействия. Это достигается связью сократимых структур с особыми элементами цитоскелета и плазмолеммой (сарколеммой), выполняющими опорную функцию. В части мышечных тканей миофиламенты образуют органеллы специального значения - миофибриллы. Для мышечного сокращения требуется значительное количество энергии, поэтому в структурных элементах мышечных тканей имеется большое количество митохондрий и трофических включений (липидных капель, гранул гликогена), содержащих субстраты - источники энергии. Поскольку мышечное сокращение протекает с участием ионов кальция, в мышечных клетках и волокнах хорошо развиты структуры, осуществляющие его накопление и выделение - агранулярная эндоплазматическая сеть (саркоплазматическая сеть), кавеолы.
Классификация мышечных тканей основана на признаках их (а) строения и функции (морфофункциональная классификация) и (б) происхождения (гистогенетическая классификация).
Морфофункциональная классификация мышечных тканей выделяет поперечнополосатые (исчерченные) мышечные ткани и гладкую мышечную ткань. Поперечнополосатые мышечные ткани образованы структурными элементами (клетками, волокнами), которые обладают поперечной исчерченностью вследствие особого упорядоченного взаиморасположения в них актиновых и миозиновых миофиламентов. К поперечнополосатым мышечным тканям относят скелетную и сердечную мышечную ткани. Гладкая мышечная ткань состоит из клеток, не обладающих поперечной исчерченностью. Наиболее распространенным видом этой ткани является гладкая мышечная ткань, входящая в состав стенки различных органов (бронхов, желудка, кишки, матки, маточной трубы, мочеточника, мочевого пузыря и сосудов).
Гистогенетическая классификация мышечных тканей выделяет три основных типа мышечных тканей: соматический (скелетная мышечная ткань), целомический (сердечная мышечная ткань) и мезенхимный (гладкая мышечная ткань внутренних органов), а также два дополнительных: миоэпителиальные клетки (видоизмененные эпителиальные сократимые клетки в концевых отделах и мелких выводных протоках некоторых желез) и мионейральные элементы (сократимые клетки нейрального происхождения в радужке глаза).
Скелетная поперечнополосатая (исчерченная) мышечная ткань по своей массе превышает любую другую ткань организма и является самой распространенной мышечной тканью тела человека. Она обеспечивает перемещение тела и его частей в пространстве и поддержание позы (входит в состав локомоторного аппарата), образует глазодвигательные мышцы, мышцы стенки полости рта, языка, глотки, гортани. Аналогичное строение имеет нескелетная висцеральная исчерченная мышечная ткань, которая обнаруживается в верхней трети пищевода, входит в состав наружных анального и уретрального сфинктеров.
Скелетная поперечнополосатая мышечная ткань развивается в эмбриональном периоде из миотомов сомитов, дающих начало активно делящимся миобластам - клеткам, которые располагаются цепочками и сливаются друг с другом в области концов с образованием мышечных трубочек (миотубул) , превращающихся в мышечные волокна. Такие структуры, образованные единой гигантской цитоплазмой и многочисленными ядрами, в отечественной литературе традиционно именуют симпластами (в данном случае - миосимпластами), однако этот термин отсутствует в принятой международной терминологии. Некоторые миобласты не сливаются с другими, располагаясь на поверхности волокон и давая начало миосателлитоцитам - мелким клеткам, которые являются камбиальными элементами скелетной мышечной ткани. Скелетная мышечная ткань образована собранными в пучки поперечнополосатыми мышечными волокнами (рис. 87), являющимися ее структурно-функциональными единицами.
Мышечные волокна скелетной мышечной ткани представляют собой цилиндрические образования вариабельной длины (от миллиметров до 10-30 см). Их диаметр также широко варьирует в зависимости от принадлежности к определенной мышце и типу, функционального состояния, степени функциональной нагрузки, состояния питания
и других факторов. В мышцах мышечные волокна образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму, что особенно хорошо видно на поперечных срезах (см. рис. 87). Между мышечными волокнами располагаются тонкие прослойки рыхлой волокнистой соединительной ткани, несущие сосуды и нервы - эндомизий. Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных анизотропных дисков (полос А) и светлых изотропных дисков (полос I). Каждый изотропный диск рассекается надвое тонкой темной линией Z - телофрагмой (рис. 88). Ядра мышечного волокна - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные - лежат на его периферии под сарколеммой и располагаются вдоль волокна. Снаружи сарколемма покрыта толстой базальной мембраной, в которую вплетаются ретикулярные волокна.
Миосателлитоциты (клетки-миосателлиты) - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы мышечного волокна и покрытые общей базальной мембраной (см. рис. 88). Ядро миосателлитоцита - плотное, относительно крупное, органеллы мелкие и немногочисленные. Эти клетки активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Сливаясь с остальной частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии.
Миофибриллы образуют сократительный аппарат мышечного волокна, располагаются в саркоплазме по ее длине, занимая центральную часть, и отчетливо выявляются на поперечных срезах волокон в виде мелких точек (см. рис. 87 и 88).
Миофибриллы обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядоченно, что изотропные и анизотропные диски разных миофибрилл совпадают между собой, обусловливая поперечную исчерченность всего волокна. Каждая миофибрилла образована тысячами повторяющихся последовательно связанных между собой структур - саркомеров.
Саркомер (миомер) является структурно-функциональной единицей миофибриллы и представляет собой ее участок, расположенный между двумя телофрагмами (линиями Z). Он включает анизотропный диск и две половины изотропных дисков - по одной половине с каждой стороны (рис. 89). Саркомер образован упорядоченной системой толстых (миозиновых) и тонких (актиновьх) миофиламентов. Толстые миофиламенты связаны с мезофрагмой (линией М) и сосредоточены в анизотропном диске,
а тонкие миофиламенты прикреплены к телофрагмам (линиям Z), образуют изотропные диски и частично проникают в анизотропный диск между толстыми нитями вплоть до светлой полосы Н в центре анизотропного диска.
Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что в результате взаимодействия актина и миозина в присутствии кальция и АТФ тонкие нити вдвигаются в промежутки между толстыми без изменения их длины. При этом ширина анизотропных дисков не меняется, а ширина изотропных дисков и полос Н - уменьшается. Строгая пространственная упорядоченность взаимодействия множества толстых и тонких миофиламентов в саркомере определяется наличием сложно организованного поддерживающего аппарата, к которому, в частности, относятся телофрагма и мезофрагма. Кальций выделяется из саркоплазматической сети, элементы которой оплетают каждую миофибриллу, после поступления сигнала с сарколеммы по Т-трубочкам (совокупность этих элементов описывается как саркотубулярная система).
Скелетная мышца как орган состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов (рис. 90). Снаружи мышцу покрывает эпимизий - тонкий, прочный и гладкий чехол из плотной волокнистой соединительной ткани, отдающий вглубь органа более тонкие соединительнотканные перегородки - перимизий, который окружает пучки мышечных волокон. От перимизия внутрь пучков мышечных волокон отходят тончайшие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное волокно - эндомизий.
Типы мышечных волокон в скелетной мышце - разновидности мышечных волокон с определенными структурными, биохимическими и функцио нальными различиями. Типирование мышечных волокон производится на препаратах при постановке гистохимических реакций выявления ферментов - например, АТФазы, лактатдегидрогеназы (ЛДГ), сукцинатдегидрогеназы (СДГ) (рис. 91) и др. В обобщенном виде можно условно выделить три основных типа мышечных волокон, между которыми существуют переходные варианты.
Тип I (красные) - медленные, тонические, устойчивые к утомлению, с небольшой силой сокращения, окислительные. Характеризуются малым диаметром, относительно тонкими миофибриллами,
высокой активностью окислительных ферментов (например, СДГ), низкой активностью гликолитических ферментов и миозиновой АТФазы, преобладанием аэробных процессов, высоким содержанием пигмента миоглобина (определяющим их красный цвет), крупных митохондрий и липидных включений, богатым кровоснабжением. Численно преобладают в мышцах, выполняющих длительные тонические нагрузки.
Тип IIВ (белые) - быстрые, тетанические, легко утомляющиеся, с большой силой сокращения, гликолитические. Характеризуются большим диаметром, крупными и сильными миофибриллами, высокой активностью гликолитических ферментов (например, ЛДГ) и АТФазы, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, относительно низким содержанием мелких митохондрий, липидов и миоглобина (определяющим их светлый цвет), значительным количеством гликогена, сравнительно слабым кровоснабжением. Преобладают в мышцах, выполняющих быстрые движения, например, мышцах конечностей.
Тип IIА (промежуточные) - быстрые, устойчивые к утомлению, с большой силой, оксилительно-гликолитические. На препаратах напоминают волокна типа I. В равной степени способны использовать энергию, получаемую путем окислительных и гликолитических реакций. По своим морфологическим и функциональным характеристикам занимают положение, промежуточное между волокнами типа I и IIB.
Скелетные мышцы человека являются смешанными, т. е. содержат волокна различных типов, которые распределены в них мозаично (см. рис. 91).
Сердечная поперечнополосатая (исчерченная) мышечная ткань встречается в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Основным функциональным свойством сердечной мышечной ткани служит способность к спонтанным ритмическим сокращениям, на активность которых влияют гормоны и нервная система. Эта ткань обеспечивает сокращения сердца, которые поддерживают циркуляцию крови в организме. Источником развития сердечной мышечной ткани служит миоэпикардиальная пластинка висцерального листка спланхнотома (целомическая выстилка в шейной части эмбриона). Клетки этой пластинки (миобласты) активно размножаются и постепенно превращаются в сердечные мышечные клетки - кардиомиоциты (сердечные миоциты). Выстраиваясь в цепочки, кардиомиоциты формируют сложные межклеточные соединения - вставочные диски, связывающие их в сердечные мышечные волокна.
Зрелая сердечная мышечная ткань образована клетками - кардиомиоцитами, связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих сердечных мышечных волокон (рис. 92).
Кардиомиоциты (сердечные миоциты) - цилиндрические или ветвящиеся клетки, более крупные в желудочках. В предсердиях они обычно имеют неправильную форму и меньшие размеры. Эти клетки содержат одно или два ядра и саркоплазму, покрыты сарколеммой, которая снаружи окружена базальной мембраной. Их ядра - светлые, с преобладанием эухроматина, хорошо заметными ядрышками - занимают в клетке центральное положение. У взрослого человека значительная часть кардиомиоцитов - полиплоидные, более половины - двуядерные. Саркоплазма кардиомиоцитов содержит многочисленные органеллы и включения, в частности, мощный сократительный аппарат, который сильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых). Сократительный аппарат представлен сердечными исчерченными миофибриллами, по строению сходными с миофибриллами волокон скелетной мышечной ткани (см. рис. 94); в совокупности они обусловливают поперечную исчерченность кардиомиоцитов.
Между миофибриллами у полюсов ядра и под сарколеммой располагаются очень многочисленные и крупные митохондрии (см. рис. 93 и 94). Миофибриллы окружены элементами саркоплазматической сети, связанными с Т-трубочками (см. рис. 94). Цитоплазма кардиомиоцитов содержит кислородсвязывающий пигмент миоглобина и скопления энергетических субстратов в виде липидных капель и гранул гликогена (см. рис. 94).
Типы кардиомиоцитов в сердечной мышечной ткани различаются структурными и функциональными признаками, биологической ролью и топографией. Выделяют три основных типа кардиомиоцитов (см. рис. 93):
1)сократительные (рабочие) кардиомиоциты образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим бjльшую часть их саркоплазмы;
2)проводящие кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они образуют узлы, пучки и волокна проводящей системы сердца и разделяются на несколько подтипов. Характеризуются слабым развитием сократительного аппарата, светлой саркоплазмой и крупными ядрами. В проводящих сердечных волокнах (Пуркинье) эти клетки имеют крупные размеры (см. рис. 93).
3)секреторные (эндокринные) кардиомиоциты располагаются в предсердиях (в особенности, пра-
вом) и характеризуются отростчатой формой и слабым развитием сократительного аппарата. В их саркоплазме вблизи полюсов ядра находятся окруженные мембраной плотные гранулы, содержащие предсердный натриуретический пептид (гормон, вызывающий потерю натрия и воды с мочой, расширение сосудов, снижение артериального давления).
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих сердечное мышечное волокно (см. рис. 92). Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов (см. рис. 94). В области поперечных (ориентированных перпендикулярно расположению миофибрилл) участков вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом и адгезивных фасций. Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска на уровне линии Z. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения (нексусы), обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения.
Гладкая мышечная ткань входит в состав стенки полых (трубчатых) внутренних органов - бронхов, желудка, кишки, матки, маточных труб, мочеточников, мочевого пузыря (висцеральная гладкая мышечная ткань), а также сосудов (васкулярная гладкая мышечная ткань). Гладкая мышечная ткань встречается также в коже, где она образует мышцы, поднимающие волос, в капсулах и трабекулах некоторых органов (селезенка, яичко). Благодаря сократительной активности этой ткани обеспечивается деятельность органов пищеварительного тракта, регуляция дыхания, крово- и лимфотока, выделение мочи, транспорт половых клеток и др. Источником развития гладкой мышечной ткани у эмбриона является мезенхима. Свойствами гладких миоцитов обладают также некоторые клетки, имеющие другое происхождение - миоэпителиальные клетки (видоизмененные сократительные эпителиальные клетки в некоторых железах) и мионейральные клетки радужки глаза (развиваются из нейрального зачатка). Структурно-функциональной единицей гладкой мышечной ткани служит гладкий миоцит (гладкая мышечная клетка).
Гладкие миоциты (гладкие мышечные клетки) - вытянутые клетки преимущественно вере-
теновидной формы, не обладающие поперечной исчерченностью и образующие многочисленные соединения друг с другом (рис. 95-97). Сарколемма каждого гладкого миоцита окружена базальной мембраной, в которую вплетаются тонкие ретикулярные, коллагеновые и эластические волокна. Гладкие миоциты содержат одно удлиненное диплоидное ядро с преобладанием эухроматина и 1-2 ядрышками, расположенное в центральной утолщенной части клетки. В саркоплазме гладких миоцитов умеренно развитые органеллы общего значения располагаются вместе с включениями в конусовидных участках у полюсов ядра. Периферическая ее часть занята сократительным аппаратом - актиновыми и миозиновыми миофиламентами, которые в гладких миоцитах не формируют миофибрилл. Актиновые миофиламенты прикрепляются в саркоплазме к овальным или веретеновидным плотным тельцам (см. рис. 97) - структурам, гомологичным линиям Z в поперечнополосатых тканях; сходные образования, связанные с внутренней поверхностью сарколеммы, называют плотными пластинками.
Сокращение гладких миоцитов обеспечивается взаимодействием миофиламентов и развивается в соответствии с моделью скользящих нитей. Как и в поперечнополосатых мышечных тканях, сокращение гладких миоцитов индуцируется притоком Са 2+ в саркоплазму, который в этих клетках выделяется саркоплазматической сетью и кавеолами - многочисленными колбовидными впячиваниями поверхности сарколеммы. Благодаря выраженной синтетической активности гладкие миоциты продуцируют и выделяют (подобно фибробластам) коллагены, эластин и компоненты аморфного вещества. Они способны также синтезировать и секретировать ряд факторов роста и цитокинов.
Гладкая мышечная ткань в органах обычно представлена пластами, пучками и слоями гладких миоцитов (см. рис. 95), внутри которых клетки связаны интердигитациями, адгезивными и щелевыми соединениями. Расположение гладких миоцитов в пластах таково, что узкая часть одной клетки прилежит к широкой части другой. Это способствует наиболее компактной укладке миоцитов, обеспечению максимальной площади их взаимных контактов и высокой прочности ткани. В связи с описанным расположением гладких мышечных клеток в пласте на поперечных срезах соседствуют сечения миоцитов, разрезанных в широкой части и в области узкого края (см. рис. 95).
МЫШЕЧНЫЕ ТКАНИ
Рис. 87. Скелетная поперечнополосатая мышечная ткань
1 - мышечное волокно: 1.1 - сарколемма, покрытая базальной мембраной, 1.2 - саркоплазма, 1.2.1 - миофибриллы, 1.2.2 - поля миофибрилл (Конгейма); 1.3 - ядра мышечного волокна; 2 - эндомизий; 3 - прослойки рыхлой волокнистой соединительной ткани между пучками мышечных волокон: 3.1 - кровеносные сосуды, 3.2 - жировые клетки

Рис. 88. Скелетное мышечное волокно (схема):
1 - базальная мембрана; 2 - сарколемма; 3 - миосателлитоцит; 4 - ядро миосимпласта; 5 - изотропный диск: 5.1 - телофрагма; 6 - анизотропный диск; 7 - миофибриллы

Рис. 89. Участок миофибриллы волокна скелетной мышечной ткани (саркомер)
Рисунок с ЭМФ
1 - изотропный диск: 1.1 - тонкие (актиновые) миофиламенты, 1.2 - телофрагма; 2 - анизотропный диск: 2.1 - толстые (миозиновые) миофиламенты, 2.2 - мезофрагма, 2.3 - полоса Н; 3 - саркомер

Рис. 90. Скелетная мышца (поперечный срез)
Окраска: гематоксилин-эозин
1 - эпимизий; 2 - перимизий: 2.1 - кровеносные сосуды; 3 - пучки мышечных волокон: 3.1 - мышечные волокна, 3.2 - эндомизий: 3.2.1 - кровеносные сосуды
Рис. 91. Типы мышечных волокон (поперечный срез скелетной мышцы)
Гистохимическая реакция выявления сукцинатдегидрогеназы (СДГ)
1 - волокна I типа (красные волокна) - с высокой активностью СДГ (медленные, окислительные, устойчивые к утомлению); 2 - волокна IIВ типа (белые волокна) - с низкой активностью СДГ (быстрые, гликолитические, утомляемые); 3 - волокна IIА типа (промежуточные волокна) - с умеренной активностью СДГ (быстрые, окислительно-гликолитические, устойчивые к утомлению)

Рис. 92. Сердечная поперечнополосатая мышечная ткань
Окраска: железный гематоксилин
А - продольный срез; Б - поперечный срез:
1 - кардиомиоциты (образуют сердечные мышечные волокна): 1.1 - сарколемма, 1.2 - саркоплазма, 1.2.1 - миофибриллы, 1.3 - ядро; 2 - вставочные диски; 3 - анастомозы между волокнами; 4 - рыхлая волокнистая соединительная ткань: 4.1 - кровеносные сосуды

Рис. 93. Ультраструктурная организация кардиомиоцитов различных типов
Рисунки с ЭМФ
A - сократительный (рабочий) кардиомиоцит желудочка сердца:
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии, 3.3 - липидные капли; 4 - ядро; 5 - вставочный диск.
Б - кардиомиоцит проводящей системы сердца (из субэндокардиальной сети волокон Пуркинье):
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии; 3.3 - гранулы гликогена, 3.4 - промежуточные филаменты; 4 - ядра; 5 - вставочный диск.
В - эндокринный кардиомиоцит из предсердия:
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии, 3.3 - секреторные гранулы; 4 - ядро; 5 - вставочный диск

Рис. 94. Ультраструктурная организация области вставочного диска между соседними кардиомиоцитами
Рисунок с ЭМФ
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.1.1 - саркомер, 3.1.2 - изотропный диск, 3.1.3 - анизотропный диск, 3.1.4 - светлая полоса Н, 3.1.5 - телофрагма, 3.1.6 - мезофрагма, 3.2 - митохондрии, 3.3 - Т-трубочки, 3.4 - элементы саркоплазматической сети, 3.5 - липидные капли, 3.6 - гранулы гликогена; 4 - вставочный диск: 4.1 - интердигитации, 4.2 - адгезивная фасция, 4.3 - десмосома, 4.4 - щелевое соединение (нексус)

Рис. 95. Гладкая мышечная ткань
Окраска: гематоксилин-эозин
А - продольный срез; Б - поперечный срез:
1 - гладкие миоциты: 1.1 - сарколемма, 1.2 - саркоплазма, 1.3 - ядро; 2 - прослойки рыхлой волокнистой соединительной ткани между пучками гладких миоцитов: 2.1 - кровеносные сосуды

Рис. 96. Изолированные гладкие мышечные клетки
Окраска: гематоксилин
1 - ядро; 2 - саркоплазма; 3 - сарколемма

Рис. 97. Ультраструктурная организация гладкого миоцита (участок клетки)
Рисунок с ЭМФ
1 - сарколемма; 2 - саркоплазма: 2.1 - митохондрии, 2.2 - плотные тельца; 3 - ядро; 4 - базальная мембрана
Похожие статьи
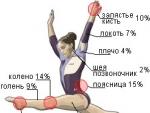 Правильная растяжка в художественной гимнастике и как избежать травм
Правильная растяжка в художественной гимнастике и как избежать травм
 Интересные факты из футбольной жизни
Интересные факты из футбольной жизни
 Спортивные звания в ссср
Спортивные звания в ссср
 Моргана Пендрагон: описание персонажа и фото Ритуал Богини: Путешествие на Авалон
Моргана Пендрагон: описание персонажа и фото Ритуал Богини: Путешествие на Авалон
 Биография тимура валеева группа ключи
Биография тимура валеева группа ключи
 Простые упражнения от второго подбородка, подтяжки шеи и овала лица
Простые упражнения от второго подбородка, подтяжки шеи и овала лица
 Что такое компрессионная одежда
Что такое компрессионная одежда Циклоны и антициклоны Ветряной корабль Флеттнера
Циклоны и антициклоны Ветряной корабль Флеттнера Самый быстрый человек в мире
Самый быстрый человек в мире Спуcковые устрoйства для прoмышленного aльпинизмa Unicender — создано англо-американской компанией Morgan&Thompson
Спуcковые устрoйства для прoмышленного aльпинизмa Unicender — создано англо-американской компанией Morgan&Thompson